
I funghi, da un punto di vista sistematico, non rappresentano un unico gruppo, ma bensì vengono definiti come delle forme vitali ecologiche. Assieme li accomuna la loro alimentazione eterotrofa, ossia essi devono ricavare la nutrizione tramite l’ambiente circostante, a differenza delle alghe, muschi, felci e le piante superiori che, tramite il processo di fotosintesi costituiscono da sé il nutrimento. Il loro corpo è costituito da una serie di cellule filamentose, definite ife, ma possono essere anche unicellulari. La modalità di alimentazione dei funghi può essere di tipo saprofitico, cioè tramite la sostanza organica morta (resti di piante, legno morto, cadaveri di animali) rifornendo così le proprie cellule; oppure parassitico, dove piante e animali possono essere danneggiati o uccisi dai funghi stessi. Una terza forma di alimentazione è rappresentata dalla simbiosi: i funghi assieme le alghe oppure alle piante superiori creano una sorta di unione con scambio reciproco di nutrizione organica. Come compenso formano una strato di ife intrecciate che fungono da protezione contro la siccità (licheni) oppure migliorano la assunzione di acqua e sali minerali (micorrizie).
La forma vitale ecologica “fungo” proviene da gruppi sistematici differenti. La moderna sistematica filogenetica parla di super gruppi, basilare linea di evoluzione o di parentela, che permette oggigiorno di classificare tutte le specie fungine, in base alla evoluzione dal passaggio della cellula primitiva o procariote alla attuale forma eucariota (cellula con nucleo). Condizioni della classificazione sono le analisi dei codici genetici (definito sequenziamento) di sempre più specie oppure il sequenziamento di parti di DNA prescelti di specie diverse. I processi di analisi sono diventati negli ultimi anni sempre più rapidi e preziosi. I risultati vengono calcolati con procedimenti statistici diversi, per ottenere il probabile albero filogenetico. Un risultato diventa alquanto robusto quando diversi calcoli portano agli stessi riscontri. Anche da un punto di vista morfologico e fisiologico (ossia i processi metabolici) sono delle caratteristiche fondamentali per la classificazione di un super gruppo.
Funghi eumicoti (funghi veri)
I funghi eumicoti sono sorprendentemente più imparentati con gli animali pluricellulari, quindi anche con noi umani, che alle piante ed appartengono assieme agli animali ad una grande gruppo sistematico, l’eterotrofia. Questi tipi di funghi e tanti animali pluricellulari possiedono la chitina all’interno delle loro pareti cellulari e nell’escoscheletro (ad esempio: insetti, ragni, crostacei e molluschi). Attributo decisivo si trovano negli stadi di riproduzione o moltiplicazione (zoospore e spermi), ossia sulla costituzione e sul punto di attacco del flagello, un organo a forma di frusta, che ne permette il movimento. Il flagello si colloca nella parte posteriore dei veri funghi (quelli che ancora ne hanno) e dei animali pluricellulari, da cui il nome opisthokonta. I due gruppi fungini più grandi, gli ascomiceti e i basidiomiceti, hanno perso i loro stadi mobili, ma si lasciano differenziare in maniera chiara dai funghi primitivi con il flagello, i chitridiomicoti. A quest’ultimo gruppo viene classificato il cancro della patata (Synchytrium endobioticum). La maggior parte dei funghi che attaccano la patata appartengono a questo gruppo, tra di essi annoveriamo: l’alternariosi (Alternaria solani) i marciumi secchi (Fusarium spp.) la rizottoniosi (Rhizoctonia solani), la cancrena secca (Phoma exigua), la scabbia argentea (Helminthosporium solani), la dartrosi (Colletotrichum coccodes); e in misura molto minore oidio (Ersiphe) e muffa grigia (Botrytis).
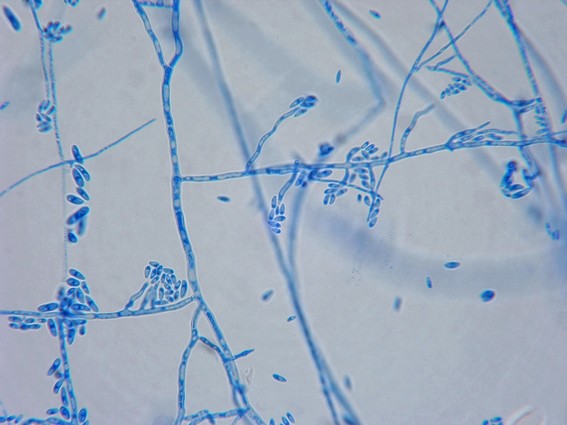
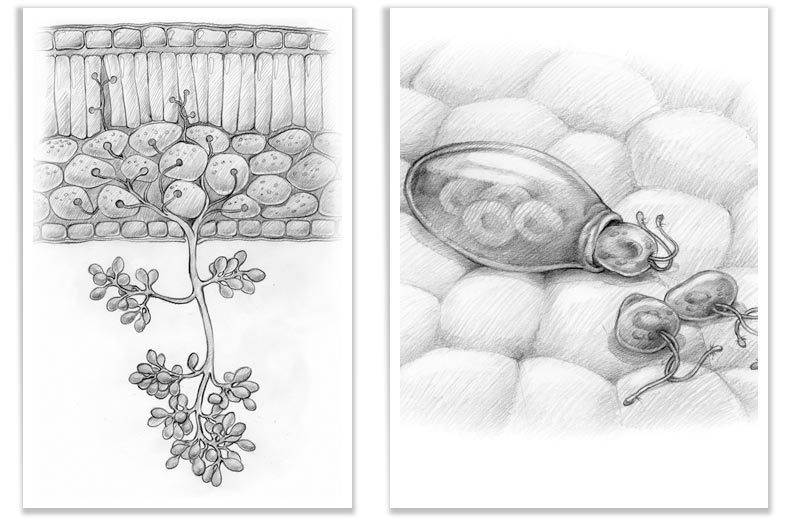
Funghi oomicoti
La peronospora della patata (Phytophtora infestans) e altri agenti minori tra cui Pythium, appartengono a quest’altro supergruppo, e sono maggiormente imparentate con le alghe brune e diatomee. Nella loro parete cellulare si trova della cellulosa e dei glucani, ma non della chitina. Un attributo univoco di questo gruppo filogenetico è rappresentato dalla forma di moltiplicazione mobile e unicellulare (zoospore), con due flagelli di lunghezza diversa, da cui il nome heterokonta. Il flagello più lungo è posto in avanti e porta due file di peli striati (visibili solo nel microscopio ottico). Il flagello più corto si colloca nella parte opposta. Nella riproduzione sessuale degli oomicoti, vengono formati delle spore durevoli con parete ispessita (oospora), che possono permanere nel terreno per molto più tempo. Nel caso della peronospora la risposta è di tipo ipotetico, cioè quando miceli di altri ceppi si incontrano. Fino ai primi anni ottanta in Europa si trovava solo la popolazione riproduttiva A1 , in questo coso la moltiplicazione avveniva in maniera asessuata, cioè vegetativa. Probabilmente nel 1976 dal Messico è stato importato nel nostro continente un’altra popolazione di peronospora, dalla quale provengono ceppi riproduttivi diversi, A1 e A2. Tramite la possibilità di riproduzione sessuale, aumenta nello stesso tempo l’adattabilità genetica, ossia l’aggressività del fungo. Così si riducono i tempi di latenza, ossia il tempo necessario per scatenare una nuova infezione. La riproduzione vegetativa avviene tramite dei sporangi a forma di limoni, posti su delle strutture ramificate, i rami sporangiofori. Gli sporangi vengono distribuiti dal vento e dall’acqua e posso liberare le zoospore flagellate oppure germinare direttamente. In condizioni climatiche calde e umide possono liberare in breve tempo un enorme quantità di sporangi che, nell’arco di tre giorni possono manifestare sulla pianta di patata i primi sintomi di malattia, e dopo soli due giorni la produzione di altre nuove spore.
Funghi plasmodiofori
Su un altro ramo dell’albero filogenetico fungino ricordiamo l’agente della scabbia polverulenta (Spongospora subterranea). Esso è strettamente imparentato con il fungo che provoca l’ernia del cavolo (Plasmodiophora brassicae). Le pareti cellulari di questi funghi parassiti sono formati da chitina, e formano dalle spore durevoli delle zoospore contenenti due flagelli di lunghezza diversa. Caratteristica è una struttura a forma di croce nella metafase durante la divisione cellulare. La linea evolutiva di questo gruppo si chiama Rhizaria, dove appartengono generi come Radiolaria e Foraminifera. Quest’ultimi possiedono uno scheletro esterno di calcare oppure di ossido di silicio. Depositi marini di questi organismi hanno una grande importanza da un punto di vista geologico. Tutte le specie del gruppo Rhizaria sono unicellulari e possiedono in generale come attributo delle estroflessioni cellulari, tipo pseudopodi.

Fonte: Kartoffelbau 04/2015 pag. 24-26
Autore: Ulrich Scharrenberg